

Как заработать свои первые деньги?
Слушайте больше на Подкасте Михалыча для молодежи
ГОУ ВПО «ЧУВАШСКИЙ ГОСУДАРСТВЕННЫЙ ПЕДАГОГИЧЕСКИЙ УНИВЕРСИТЕТ ИМ. И. Я. ЯКОВЛЕВА»
Кафедра анатомии, физиологии и гигиены детей
,
«Молекулярные основы наследственности. Строение и функции нуклеиновых кислот»
«Организация генома»
Учебное пособие по основам генетики
Чебоксары 2005
, . Учебное пособие по основам генетики. Тема: «Молекулярные основы наследственности. Строение и функции нуклеиновых кислот» «Организация генома» - Чебоксары, 2005. – 20 С.
Учебное пособие включает материалы по типам и строению нуклеиновых кислот, а также полинуклеотидных цепей ДНК и РНК, материалы по записи генетической информации в молекуле ДНК, свойствам генетического кода. Уделено внимание структурно-функциональной единице наследственной информации - гене и организации генома. Руководство предназначено для студентов, обучающихся по специальностям «Логопедия» и «Дефектология».
Научный редактор – доктор биологических наук, профессор
Рецензенты:
, доктор медицинских наук, профессор кафедры профилактической медицины ЧГУ им. ;
, кандидат биологических наук, ст. преподаватель кафедры анатомии, физиологии и гигиены детей ЧГПУ им.
© , , 2005
© ГОУ ВПО «Чувашский государственный
педагогический университет им. », 2005
Введение
Настоящее руководство посвящено разделу генетики - молекулярным основам наследственности. Как показал опыт преподавания дисциплин по основам генетики, изучение данного раздела имеет важное значение для понимания наследования признаков у организмов и закономерностей функционирования отдельных клеток и клеточных систем – тканей и органов. Естественно, что данное пособие должно стать ориентиром для дальнейшего глубокого освоения основ наследственности, строения генов, генома человека и наследственной патологии.
Чтобы стать опытным педагогом в области дефектологии и раскрыть личностные качества ребенка, ограниченные тем или иным заболеванием, формируемым за счет генетической программы, полученной от родителей и среды, в которой он воспитывается, необходимо знать основы генетики, и в частности, молекулярные основы наследственности. В дальнейшей работе эти знания помогут педагогу формировать индивидуализированную среду, ориентированную на уникальность генетической конституции каждого ребенка.
«Молекулярные основы наследственности. Строение и функции нуклеиновых кислот»
Свойство организмов передавать потомству свои признаки и особенности развития, основано на передаче из поколения в поколение наследственной информации. В середине XIX в. было установлено, что способность к наследованию признаков определяется материалом, находящимся в ядре клетки. Саттон и Т. Бовери предположили, что хромосомы являются носителями наследственной информации. В 1869 г. швейцарский химик Ф. Мишер обнаружил в клеточном ядре особое вещество кислого характера, названное им нуклеином. Это событие расценивается как открытие нуклеиновых кислот. Потребовалось несколько десятков лет, чтобы убедиться, что материальным носителем наследственной информации является только одна из составных частей хромосомы – молекула дезоксирибонуклеиновой кислоты (ДНК).
Типы нуклеиновых кислот
Термин нуклеиновые кислоты был введен в 1889 г., а в 1891 г. немецкий биохимик А. Кессель описал гидролиз нуклеиновой кислоты и установил, что она состоит из остатков сахара (пептозы), фосфорной кислоты и четырех гетероциклических оснований (пурины и пиримидины).
Особенности химического строения нуклеиновых кислот обеспечивают возможность хранения, переноса и передачи по наследству дочерним клеткам информации о структуре белковых молекул, которые синтезируются в каждой ткани на определенном этапе индивидуального развития.
Важное условие нормальной жизнедеятельности клетки и целого организма – стабильность нуклеиновых кислот. Изменения строения нуклеиновых кислот влекут за собой изменения структуры клеток или активности физиологических процессов в них, влияя на жизнеспособность клеток, тканей и организма в целом.
Структуру нуклеиновых кислот впервые установили американский биохимик Дж. Уотсон и английский физик Ф. Крик. Ее изучение имеет важное значение для понимания наследования признаков у организмов и закономерностей функционирования отдельных клеток и клеточных систем – тканей и органов.
Существуют два различных типа нуклеиновых кислот: дезоксирибонуклеиновая кислота (ДНК) и рибонуклеиновые кислоты (РНК).
ДНК – дезоксирибонуклеиновая кислота
В начале 50-х гг. было получено множество данных (большое количество экспериментов было проведено на вирусах), свидетельствующих об универсальности ДНК как носителя генетической информации. ДНК представляет собой генетический материал большинства организмов. В прокариотипических клетках, кроме основной хромосомной ДНК, часто встречаются внехромосомные ДНК - плазмиды. В эукариотических клетках основная масса ДНК расположена в клеточном ядре, где она связана с различными белками в хромосомах, а также содержится в органеллах – митохондриях и пластидах.
ДНК – это линейный, нерегулярный биологический полимер, состоящий из двух полинуклеотидных цепей, соединенных друг с другом. Пространственная конфигурация молекул ДНК была установлена в 1953 г. (модель Дж. Уотсона и Ф. Крика). Это открытие было удостоено Нобелевской премии.
Мономеры, составляющие каждую из цепей ДНК, - сложные органические соединения – нуклеотиды. Одни из важнейших компонентов нуклеотидов представляют собой гетероциклические азотистые основания, производные пирамидина или пурина.
В подавляющем большинстве случаев в состав нуклеотидов ДНК входят тимин (Т) и цитозин (Ц) – производные пирамидина, а также аденин (А) и гуанин (Г), относящиеся к производным пурина (рис. 1). Кроме того, нуклеотиды включают пятиатомный сахар (пентозу) - дезоксирибозу, а также остаток фосфорной кислоты (рис. 2).
|
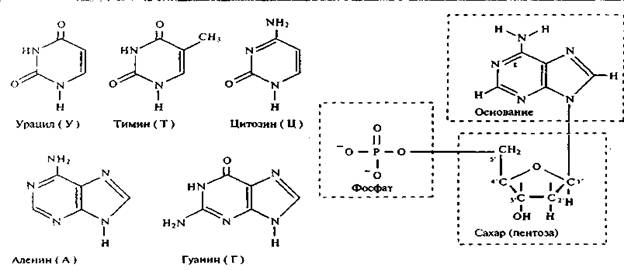
Рис. 1 Азотистые основания, входящие Рис.2. Схема строений нуклеотида
в состав нуклеотидов ДНК
ДНК – полимер с очень большой молекулярной массой; в одну молекулу может входить 108 и более нуклеотидов. В каждой полинуклеотидной цепи нуклеотиды соединяются между собой благодаря образованию фосфоэфирных связей между дезоксирибозой одного и остатком фосфорной кислоты последующего нуклеотида. При этом в начале молекулы – у первого нуклеотида – свободным от образования эфирной связи остается остаток фосфорной кислоты. Это так называемый 5 - конец молекулы. На другом конце молекулы, незадействованным в образовании фосфоэфирной связи, оказывается 3 - атом углерода дезоксирибозы - 3 - конец полинуклеотидной цепи.

![]() Объединяются две полинуклеотидные цепи в единую молекулу при помощи водородных связей, возникающих между азотистыми основаниями, входящими в состав нуклеотидов, которые образуют разные цепи. Количество таких связей между разными азотистыми основаниями неодинаковое, и вследствие этого они могут соединяться только попарно: азотистое основание А одной цепи полинуклеотидов всегда связано двумя водородными связями с Т другой цепи, а Г – тремя водородными связями с азотистым основанием Ц противоположной полинуклеотидной цепочки. Такая способность к избирательному соединению нуклеотидов, в результате чего формируются пары А-Т и Г-Ц, называется комплементарностью (рис. 3). Если известна последовательность оснований в одной цепи (пример: – Т-Ц-А-Т-Г), то благодаря принципу комплементарности легко определить последовательность оснований противоположной цепи (А-Г-Т-А-Ц).
Объединяются две полинуклеотидные цепи в единую молекулу при помощи водородных связей, возникающих между азотистыми основаниями, входящими в состав нуклеотидов, которые образуют разные цепи. Количество таких связей между разными азотистыми основаниями неодинаковое, и вследствие этого они могут соединяться только попарно: азотистое основание А одной цепи полинуклеотидов всегда связано двумя водородными связями с Т другой цепи, а Г – тремя водородными связями с азотистым основанием Ц противоположной полинуклеотидной цепочки. Такая способность к избирательному соединению нуклеотидов, в результате чего формируются пары А-Т и Г-Ц, называется комплементарностью (рис. 3). Если известна последовательность оснований в одной цепи (пример: – Т-Ц-А-Т-Г), то благодаря принципу комплементарности легко определить последовательность оснований противоположной цепи (А-Г-Т-А-Ц).
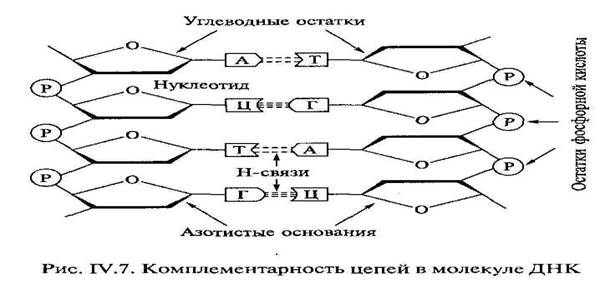 |
Рис. 3. Комплементарное соединение полинуклеотидных цепей в молекуле ДНК
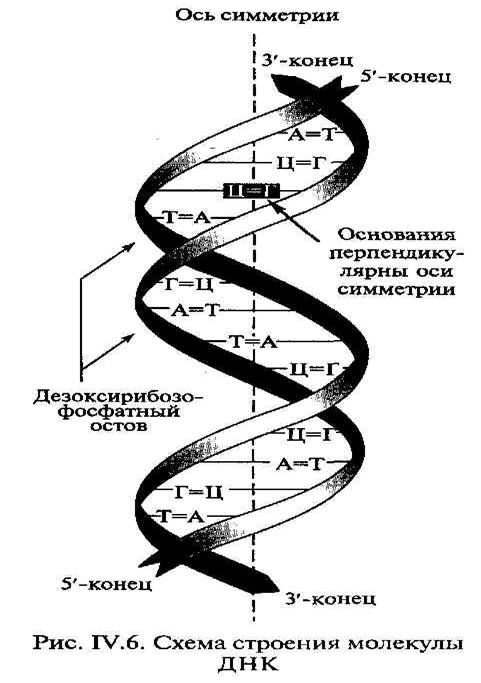
Цепи нуклеотидов образуют правозакрученные объемные спирали по 10 пар оснований в каждом витке. Последовательность соединения нуклеотидов одной цепи противоположна таковой в другой, т. е. цепи, составляющие одну молекулу ДНК, разнонаправлены (антипараллельны). Сахарофосфатные группировки нуклеотидов находятся снаружи, а комплементарно связанные нуклеотиды – внутри. Цепи закручиваются вокруг друг друга, а также вокруг общей оси и образуют двойную спираль (рис. 4).
Такая структура молекулы поддерживается в основном водородными связями. При соединении с определенными белками – гистонами – степень спирализации молекулы повышается. Молекула утолщается и укорачивается (рис. 5 Б), возникает нуклеосомная нить, представляющая собой дезоксинуклеопротеид.
 |
Последняя в результате дальнейшей спирализации образует петлистую структуру, молекула еще более укорачивается и утолщается (рис. 5 Г). Наконец, спирализация достигает максимума, возникает спираль еще более высокого уровня – суперспираль. При этом молекула ДНК, связанная с различными белками, становится различима в световой микроскоп как вытянутое, хорошо окрашиваемое тельце – хромосома (рис. 5 Д, Е).
 Рис. 4. Двойная спираль ДНК
Рис. 4. Двойная спираль ДНК
Хромосомой можно назвать самостоятельное ядерное тельце вытянутой формы, имеющее плечи и центромеру и состоящую после редупликации из двух хроматид молекул ДНК, связанных в области первичной перетяжки – центромеры. Важно отметить, что наблюдать хромосому в состоянии суперспирализации ДНК можно лишь в метафазе митоза или делений мейоза. В другие периоды жизненного цикла клетки хромосомный материал - молекулы ДНК - находятся в состоянии меньшей спирализации или деспирализованы – раскручены.
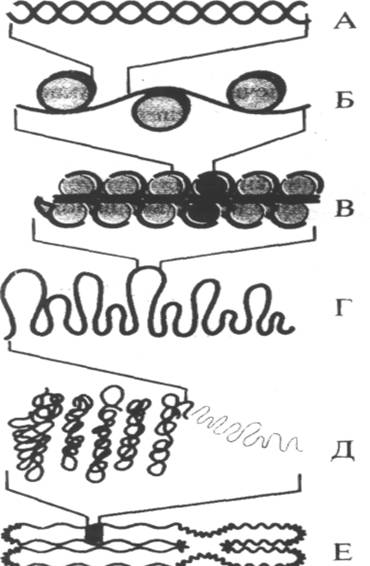
Рис. 5. Уровни спирализоции наследственного материала.
А — Короткий участок двойной спирали ДНК.
Б — Хроматин в форме «бусин на нити».
В — Хроматиновая фибрилла 30 нм, состоящая из упакованных нуклеосом.
Г — Часть хромосомы в растянутом виде.
Д — Конденсированный участок метафазной хромосомы.
Е — Целая метафазная хромосома
РНК – рибонуклеиновые кислоты
РНК так же, как ДНК, представляет собой полимер, мономерами которого являются нуклеотиды. Азотистые основания трех нуклеотидов те же самые, что входят в состав ДНК (аденин, гуанин, цитозин), четвертое — урацил — присутствует в молекуле РНК вместо тимина (см. рис. 3). Нуклеотиды РНК отличаются от нуклеотидов ДНК и по строению входящего в их состав углевода: они включают другую пентозу - рибозу (вместо дезоксирибозы). В цепочку РНК нуклеотиды соединяются благодаря образованию фосфоэфирных связей между рибозой одного и остатком фосфорной кислоты последующего нуклеотида.
РНК переносят информацию о последовательности аминокислот в белках, т. е. о структуре белков, от хромосом к месту их синтеза, и участвуют в синтезе белков. По структуре различают двухцепочечные и одноцепочечные РНК. Двухцепочечные РНК — хранители генетической информации у ряда вирусов, т. е. выполняют у них функции хромосом (так называемые РНК-содержащие вирусы).
Существует несколько видов одноцепочечных РНК. Их названия обусловлены выполняемой функцией или местонахождением в клетке.
Большую часть цитоплазмы (до 80—90%) составляет рибосомалъная РНК (рРНК), содержащаяся в рибосомах. Молекулы рРНК относительно невелики и состоят из 3— 5 тыс. нуклеотидов.
Другой вид - информационные (иРНК), или матричные РНК (мРНК), переносящие к рибосомам информацию о последовательности аминокислот в белках, которые должны синтезироваться. Размер этих РНК зависит от длины участка ДНК, на котором они синтезированы. Молекулы иРНК состоят из нуклеотидов.
Транспортные РНК (тРНК) включают 76—85 нуклеотидов и выполняют несколько функций. Они доставляют аминокислоты к месту синтеза белка, «узнают» (по принципу комплементарности) триплет иРНК, соответствующий переносимой аминокислоте, осуществляют точную ориентацию аминокислоты на рибосоме.
Запись генетической информации в молекуле ДНК
Все многообразие жизни обусловливается разнообразием белковых молекул, выполняющих в клетках различные биологические функции. Структура белков определяется набором и порядком расположения аминокислот в полипептидных цепях. Именно эта последовательность аминокислот в пептидах зашифрована в молекулах ДНК с помощью генетического кода (рис.6).
В 1954 г. Г. Гамов высказал предположение, что кодирование информации в молекулах ДНК должно осуществляться сочетаниями нескольких нуклеотидов. В многообразии белков, существующих в природе, было обнаружено около 20 различных аминокислот. Для шифровки такого их числа достаточное количество сочетаний нуклеотидов может обеспечить лишь триплетный код, в котором каждая аминокислота шифруется тремя расположенными друг за другом в полинуклеотидной цепи нуклеотидами. В этом случае комбинация из четырех нуклеотидов образует 64 триплета (43 = 64).
Второе основание
А | Г | Т | Ц | ||||
Первое основание | А | ААА ААГ Фен ААТ ААЦ Лей | АГА АГГ Сер АГТ АГЦ | АТА АТГ Тир АТТ АТЦ Стоп | АЦА АЦГ Цис АЦТ Стоп АЦЦ Три | А Г Т Ц | Третье основание |
Г | ГАА ГАГ Лей ГАТ ГАЦ | ГГА ГГГ Про ГГТ ГГЦ | ГТА ГТГ Гис ГТТ ГТЦ Глн | ГЦА ГЦГ Арг ГЦТ ГЦЦ | А Г Т Ц | ||
Т | ТАА ТАГ Илей ТАТ ТАЦ Мет | ТГА ТГГ Тре ТГТ ТГЦ | ТТА ТТГ Асн ТТТ ТТЦ Лиз | ТЦА ТЦГ Сер ТЦТ ТЦЦ Арг | А Г Т Ц | ||
Ц | ЦАА ЦАГ Вал ЦАТ ЦАЦ | ЦГА ЦГГ Ала ЦГТ ЦГЦ | ЦТА ЦТГ Асн ЦТТ ЦТЦ Глу | ЦЦА ЦЦГ Гли ЦЦТ ЦЦЦ | А Г Т Ц |
Рис.6. Таблица генетического кода в последоватепьности нуклеотидов ДНК.
Генетический код в последовательности кодонов ДНК: Фен — фенилаланин, Илей — изолейцин, Мет — метионин. Вал — валин, Тир — тирозин, Гис — гистидин, Глм — глутаминовая кислота, Лиз — лизин, Асн — аспарагин, Глу — гпутамин, Цис — цистеин. Три — триптофан, Арг— аргинин, Сер — серин, Гли — глицин. Про — пролин, Тре— треонин. Ала — аланин. Стоп — конец синтеза определенного белка
Одним из самых важных этапов в изучении функции нуклеиновых кислот стала расшифровка способа записи информации в ДНК и принцип передачи ее на белковую структуру, т. е. формулирование, генетического кода. В 1961 г. Ф. Крик и С. Бреннер показали, что каждой аминокислоте в белке соответствует триплет нуклеотидов. К этому времени уже были обнаружены транспортные РНК ( и К. Огата, 1957) и выяснена их функция, основанная на взаимодействии антикодонов и кодонов (П. Замечник, 1957). Сам же генетический код, состоящий из 64 кодонов, был установлен в 1966 г. благодаря работам М. Ниренберга, Г. Кораны и С. Очоа.
Генетическим кодом называют принцип записи наследственной информации, который состоит в том, что генетическая информация о структуре рибонуклеиновых кислот и белков заключена в ДНК в последовательности нуклеотидов одной из ее цепей. Эта цепь получила название кодогенной, а комплементарная ей цепь нуклеотидов - матричной, на ней по принципу комплементарности синтезируются молекулы РНК ![]()
![]()

(рис.7).
![]() Рис. 7. Принципы записи генетической информации
Рис. 7. Принципы записи генетической информации
Оказалось, что из 64 возможных триплетов ДНК 61 триплет кодирует различные аминокислоты; оставшиеся 3 получили название бессмысленных, или нонсенс-триплетов. Они не шифруют аминокислоты и выполняют функцию знаков препинания (стоп-триплеты) при считывании наследственной информации. К ним относятся триплеты АТТ, АЦТ, АТЦ. Кроме этого, метиониновый кодон ТАЦ является еще и стартовым триплетом, с которого начинается любой ген. Впоследствии при доработке белковой молекулы первая аминокислота метионин удаляется из полипептидной цепи.
Свойства генетического кода
Обращает на себя внимание явная избыточность кода, проявляющаяся в том, что многие аминокислоты шифруются несколькими триплетами (см. рис.6).
Это свойство триплетного кода, названное вырожденностью, имеет важное значение, так как возникновение в структуре молекулы ДНК изменений по типу замены одного нуклеотида в полинуклеотидной цепи может не изменить смысла триплета. Возникшее таким образом новое сочетание из трех нуклеотидов кодирует ту же самую аминокислоту.
В процессе изучения свойств генетического кода была обнаружена его специфичность: каждый триплет способен кодировать только одну определенную аминокислоту.
Установлено полное соответствие, идентичность кода у различных видов живых организмов. Такая универсальность генетического кода свидетельствует о единстве происхождения всего многообразия живых форм на Земле в процессе биологической эволюции.
Незначительные отличия генетического кода обнаружены в ДНК митохондрий некоторых видов. Это не противоречит в целом положению об универсальности кода, но свидетельствует в пользу определенной дивергентности в его эволюции на ранних этапах существования жизни. Расшифровка кода ДНК митохондрий различных видов показала, что во всех случаях в митохондриальных ДНК отмечается общая особенность: триплет АЦТ читается как АЦЦ, и поэтому из нонсенс-триплета он превращается в шифр аминокислоты триптофана.
Другие особенности специфичны для различных видов организмов. У дрожжей триплет ГАТ и, возможно, все семейство ГА кодирует вместо аминокислоты лейцина — треонин. У млекопитающих триплет ТАГ имеет то же значение, что и ТАЦ, и кодирует аминокислоту метионин вместо изолейцина. Триплеты ТЦГ и ТЦЦ в ДНК митохондрий некоторых видов не кодируют аминокислоты, являясь нонсенс-триплетами.
Наряду с триплетностью, вырожденностью, специфичностью и универсальностью важнейшими характеристиками генетического кода являются его непрерывность и неперекрываемость кодонов при считывании. Это означает, что последовательность нуклеотидов считывается триплет за триплетом без пропусков, при этом соседние триплеты не перекрывают друг друга, т. е. каждый отдельный нуклеотид входит в состав только одного триплета при заданной рамке считывания (рис.8).
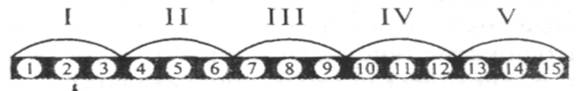
Рис. 8. Непрерывность и неперекрываемость генетического кода при считывании наследственной информации. Арабскими цифрами обозначены нуклеотиды, римскими — кодоны
Говоря о генетическом коде, мы имели в виду кодирующую цепь ДНК. Такая же последовательность нуклеотидов и в информационной, или матричной, РНК (м РНК) с учетом замены в полипептидной цепи РНК нуклеотида с азотистым основанием тимин на рибозосодержащий нуклеотид, включающий урацил — У (см. рис. 6).
Триплеты мРНК, соответствующие триплетам ДНК, также называются кодонами. В действительности именно их линейное расположение непосредственно определяют порядок включения аминокислот в синтезируемую на рибосоме полипептидную цепь.
Структурно-функциональная единица наследственной
информации — ген
Ген является элементарной единицей наследственности. Термин «ген» был предложен в 1909 г. В. Иогансеном для обозначения материальной единицы наследственности, выделенной Г. Менделем. Ген, с молекулярно-биологической точки зрения, - участок молекулы ДНК, последовательность нуклеотидов (кодонов) которого определяет последовательность нуклеотидов в одной молекуле РНК или аминокислот в одном полипептиде. В данном случае полипептид - элементарный наипростейший признак. Однако мы знаем, что многие функционально активные белки, имеющие четвертичную структурную организацию, состоят из нескольких, часто различающихся субъединиц — полипептидов. Например, гемоглобин включает две альфа-цепи и пару бета-цепей. Следовательно, за развитие такого более сложного признака отвечает уже не один, а два гена, один из которых определяет структуру альфа-цепей, а другой - бета-цепей гемоглобина. Рассматривая более сложные признаки, мы понимаем, что в их развитии принимает участие гораздо большее количество генов.
Классификация генов
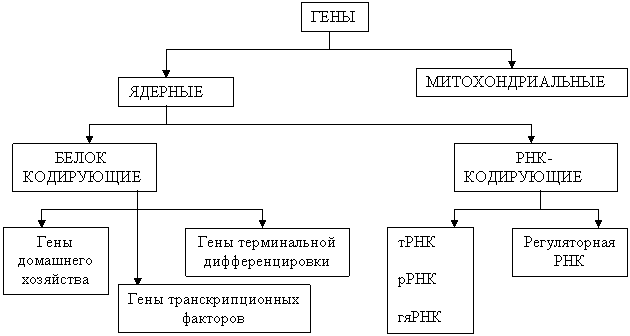
В зависимости от локализации генов в структурах клетки различают ядерные и митохондриальные гены.
По функциональному назначению гены делят на две группы: гены, кодирующие белки и гены, контролирующие синтез РНК.
Среди генов, кодирующих белки, различают:
- гены «домашнего хозяйства», продукты которых необходимы для обеспечения функции любого типа клеток;
- гены терминальной дифференцировки, обеспечивающие специализированные функции клеток;
- гены транскрипционных факторов, контролирующие особые ядерные белки, способные соединяться с регуляторными областями многих структурных генов, вызывая или активацию, или подавление транскрипции.
РНК – кодирующие гены определяют синтез различных видов РНК, обеспечения процессов трансляции, сплайсинга, синтеза молекул РНК, влияющих на функционирование других генов (регуляторное действие).
Ген представляет собой функционально прерывистую последовательность нуклеотидов, где относительно короткие кодирующие последовательности оснований чередуются с длинными некодирующими последовательностями. Экзоны являются кодирующими участками гена, контролирующими аминокислотную последовательность белков. Экзоны разделены некодирующими участками – интронами, которые вырезаются (сплайсинг) в процессе созревания иРНК и не участвуют в процессе трансляции. В настоящее время в понятие «ген» включаются не только транскрибируемые области, но и фланкирующие ген последовательности. Эти области консервативны, характеризуются постоянством нуклеотидной последовательности, необходимой для правильной работы, например: промоторная область в начале 5 ´ – области или хвостовая нетранслируемая область поли-А, расположенная на 3 ´ – конце гена, ТАТА – бокс.
Размеры генов человека могут колебаться от нескольких десятков пар нуклеотидов до многих тысяч и даже миллионов.
В 1920 г. для описания совокупности генов, заключенных в гаплоидном наборе хромосом организмов одного вида, Г. Винклер предложил термин геном. Первоначальный смысл этого термина указывал на то, что понятие генома, в отличие от генотипа, — генетическая характеристика вида в целом, а не отдельной особи. С развитием молекулярной генетики значение данного термина изменилось.
Сейчас известно, что ДНК - носитель генетической информации у большинства организмов и, следовательно, основа генома - включает в себя не только гены в современном смысле данного слова. Большая часть ДНК эукариотических клеток представлена некодирующими («избыточными») последовательностями нуклеотидов, которые не заключают в себе информации о белках и рибонуклеиновых кислотах. Таким образом, основную часть генома любого организма составляет вся ДНК его гаплоидного набора хромосом.
Однако генетическую информацию в клетках содержат не только хромосомы ядра. Жизненно важная генетическая информация бывает заключена и во внехромосомных молекулах ДНК. У бактерий к таким ДНК относятся плазмиды, в клетках эукариот - ДНК митохондрий, хлоропластов и других пластид.
В процессе онтогенеза соматические клетки могут утрачивать часть генетической информации, полученной с половыми клетками родителей, амплифицировать (избирательно редуплицировать) группы генов и значительно перестраивать исходные гены, как это происходит в лейкоцитах с генами, определяющими структуру иммуноглобулинов.
Следовательно, под геномом организма в настоящее время понимают суммарную ДНК гаплоидного набора хромосом и каждого из внехромосомных генетических элементов, содержащуюся в отдельной клетке зародышевой линии — половой клетке многоклеточного организма. Однако сформулировать определение генома отдельного биологического вида в целом непросто. В таком определении необходимо учитывать, прежде всего, генетические различия, связанные с полом организма, поскольку мужские и женские половые хромосомы различаются. Другая сложность заключается в наличии громадного числа аллельных вариантов генов и сопутствующих последовательностей, которые присутствуют в генофонде популяции и вида в целом. Вот почему, говорят лишь об усредненном геноме, который сам по себе может обладать существенными отличиями от конкретных геномов отдельных особей. Размеры геномов организмов различных видов значительно отличаются друг от друга. Причем часто не наблюдается корреляции между уровнем эволюционной сложности биологического вида и размером его генома.
Суммарное количество ДНК в гаплоидном геноме принято обозначать латинским символом С. В 1978 г. Т. Кавалье - Смит описал в качестве парадокса наблюдение того, что у эукариот транскрибируется лишь незначительная часть последовательностей нуклеотидов генома (у человека около 3% генома). В соответствии с этим необъясненный до недавнего времени феномен значительной избыточности генома эукариот в отношении некодирующих последовательностей нуклеотидов известен в генетике под названием парадокса С.
Организация генома
Структурная организация генома — фундаментальный таксономический признак, лежащий в основе современной систематики всего живого мира. В соответствии со структурной организацией генома все живые организмы разделяют на: прокариот и эукариот.
К прокариотам относят организмы, геном которых не заключен в ядро, ограниченное оболочкой, и представляет собой одну кольцевую молекулу ДНК, а также ДНК плазмид, не образующих связей с гистонами и другими белками. Надцарство прокариот включает в себя четыре группы организмов: архебактерий, эубактерий, цианобактерий и микоплазм.
Клетки эукариот содержат оформленное ядро, и их генетический материал представлен линейными молекулами ДНК — хромосомами, причем каждая из них у большинства эукариот имеет пару.
Характерная структура генома прокариот и эукариот накладывает отпечаток на морфологические, физиологические и биохимические особенности этих организмов, которые изначально определяются генетической информацией, реализующейся через экспрессию - активацию соответствующих генов.
Генетическая информация о структуре отдельных белков и нуклеиновых кислот у всех организмов заключена в молекулах ДНК или РНК в виде последовательностей нуклеотидов, называемых генами. Однако одной информации о структуре макромолекул, кодируемых генами, недостаточно для их функционирования. Координированная работа генов возможна лишь благодаря наличию тонких регуляторных механизмов, определяющих место и время экспрессии конкретного гена или группы генов. Для того, чтобы экспрессия гена была регулируемой, он должен содержать индивидуальную регуляторную метку, по которой регулирующие системы клетки или организма могли бы оказать на него необходимое воздействие. В соответствии с этим любой ген состоит из двух основных функциональных частей — последовательностей нуклеотидов — регуляторной и структурной.
Регуляторная часть обеспечивает первые этапы реализации генетической информации, заключенной в структурной части гена, которая, в свою очередь, содержит информацию о структуре конкретных нуклеиновых кислот или белков. Поэтому размер гена складывается из размеров его структурной и регуляторной частей. Однако определить протяженность гена не так просто, особенно в случае генов эукариот.
Отдельные элементы регуляторной области генов, называемые энхансерами, могут располагаться перед структурной частью гена, позади нее или даже в ней самой. В структурной части большинства эукариотических генов кодирующие последовательности нуклеотидов — экзоны перемежаются протяженными некодирующими последовательностями — интронами. Суммарный размер интронов, как правило, многократно превышает общий размер экзонов гена. Исходя из этого факта, можно сделать вывод о том, что геном любого эукариотического организма содержит не только последовательность нуклеотидов с генетической информацией о белках и рибонуклеиновых кислотах (тРНК, рРНК и некоторых других), но и большое количество последовательностей нуклеотидов, не несущих информации.
Помимо интронов, в геноме эукариот имеется большое количество других некодирующих последовательностей нуклеотидов, главным образом различных повторяющихся последовательностей. Поэтому общая длина некодирующих последовательностей нуклеотидов в геноме эукариот в десятки раз превышает длину кодирующих последовательностей.
Как у прокариотических, так и у эукариотических организмов все гены располагаются линейно, друг за другом, в составе отдельных молекул ДНК - хромосом.
У диплоидных организмов, клетки которых содержат по одному набору хромосом каждого из родителей, одинаковые хромосомы получили название гомологичных хромосом. Гомологичными называются хромосомы, одинаковые по строению, несущие в одних и тех же местах — локусах аллелъные гены, которые отвечают за развитие одних и тех же признаков. Гомологичными являются и одинаковые хромосомы разных организмов одного биологического вида. Количество хромосом в гаплоидном наборе принято обозначать латинской буквой «n». В соматических клетках каждая хромосома имеет пару, и поэтому диплоидный набор хромосом обозначается как 2п.
Гены и некодирующие последовательности нуклеотидов, заключенные в хромосомах, представляют большую часть генома организма. Зрелые клетки зародышевой линии (гаметы - яйцеклетки, сперматозоиды) многоклеточных организмов содержат по одному (гаплоидному) набору хромосом, т. е. в них находится одна хромосома из каждой пары гомологичных хромосом. Кроме того, здесь же имеются и внехромосомные генетические элементы.
Поскольку гены, входящие в состав отдельных хромосом, находятся в одной молекуле ДНК, они образуют отдельную группу сцепления и в отсутствие рекомбинации, например, в результате кроссинговера вместе передаются дочерним клеткам. Остаются до конца непонятыми физиологическое значение распределения генов по отдельным хромосомам и природа факторов, определяющих число хромосом в геноме эукариот. Например, невозможно объяснить эволюционные механизмы появления большого числа хромосом у конкретных организмов только ограничениями, накладываемыми на максимальный размер молекул ДНК, входящих в состав этих хромосом. Так, геном американской амфибии Аmphiuma содержит почти в 30 раз больше ДНК, чем геном человека, и вся она заключена только в 28 хромосомах. Однако даже самая маленькая из них больше самой крупной хромосомы человека. Неизвестны пока и факторы, ограничивающие верхний предел числа хромосом у эукариот.
По-видимому, большинство особенностей структурной и функциональной организации генома, в том числе наличие множества неинформативных последовательностей нуклеотидов, должны обеспечивать надежность его функционирования. Это обусловливает целостность передачи генетического материала от родительских клеток дочерним на протяжении многих клеточных поколений и корректную работу генов, проявляющуюся в безошибочной и упорядоченной во времени экспрессии генов.
В отличие от эукариот геном прокариот гораздо лаконичнее. Количество некодирующих последовательностей нуклеотидов минимально. Многие механизмы регуляции экспрессии генов, использующиеся у эукариот, никогда не встречаются у прокариот. Таким образом, простота строения генома прокариот объясняется, прежде всего, их упрощенным жизненным циклом, на протяжении которого прокариотические клетки, как правило, не претерпевают сложных морфофизиологических преобразований, связанных с переключением экспрессии одних групп генов на другие, что имеет место в онтогенезе эукариот.
Как уже упоминалось выше, в отличие от прокариот основная часть генома эукариот находится в ядре. Так же, как и у прокариот, информационной макромолекулой генома эукариот является ДНК, которая неравномерно распределена по нескольким хромосомам в виде комплексов с многочисленными белками. Эти ДНК-белковые комплексы неделящейся клетки эукариот получили название хроматина.
На протяжении клеточного цикла хроматин претерпевает высокоупорядоченные структурные преобразования в виде последовательных конденсаций —деконденсаций.
В соматических клетках при максимальной конденсации в метафазе митоза эти преобразования сопровождаются формированием видимых в микроскопе метафазных хромосом. Как морфология метафазных хромосом, так и их число — уникальные характеристики вида. Совокупность качественных и количественных признаков хромосомного набора соматических клеток эукариот получила название кариотипа. Эти признаки (цитогенетические) широко используются в биологической систематике и представляют собой один из критериев вида.
Таким образом, геном эукариот существенно отличается от генома прокариот по ряду признаков, среди которых необходимо отметить его избыточность. Содержание ДНК у эукариот в расчете на одну клетку в среднем на два-три порядка выше, чем у прокариот.
Говоря о генетическом коде, мы имели в виду матричную цепь ДНК. Такая же последовательность нуклеотидов и в информационной, или матричной, РНК (мРНК) с учетом комплементарности и замены в полипептидной цепи РНК нуклеотида с азотистым основанием тимин на рибозосодержащий нуклеотид, включающий урацил.
Триплеты мРНК, соответствующие триплетам ДНК, также называются кодонами. В действительности именно они непосредственно определяют последовательность включения аминокислот в синтезируемую на рибосоме полипептидную цепь. Это последовательность из трех нуклеотидов в молекуле мРНК, обеспечивающих включение одного аминокислотного остатка в полипептидную цепь, либо сигнал начала или завершения транскрипции или трансляции.
Таким образом, геном - это полная генетическая система клетки, определяющая характер индивидуального развития организма и наследственную передачу в ряду поколений всех структурных и функциональных признаков.
Заключение
Вы ознакомились с основными аспектами молекулярных основ наследственности и организацией генома человека.
Свойство организмов воспроизводить себе подобных, т. е. передавать потомкам свои признаки и особенности развития, основано на передаче из поколения в поколение наследственной информации. В настоящее время не вызывает сомнений тот факт, что организм является результатом сложного взаимодействия генетической программы, унаследованной от родителей и постоянно меняющихся условий внешней среды. Наследственная информация, реализующаяся в процессе индивидуального развития организма, обеспечивает формирование всех признаков и свойств, но не как стабильных, а как способных к определенной изменчивости. В генетической конституции организма заложена способность сохранять постоянство внутренней среды при колеблющихся внешних условиях. Среда воздействует на тот или иной материальный субстрат, являющийся результатом действия генов. Вне явления наследственности невозможны никакие процессы жизнедеятельности клетки, развития особи и эволюции организмов.
Социальная природа человека во многом становится определяющим фактором в реализации патологических генотипов. Проведение медико-генетических мероприятий, направленных на предотвращение влияния вредоносных факторов внешней среды, создание условий, способствующих реализации нормальных генотипов и препятствующих развитию паталогических, терапия ряда наследственных болезней способны уменьшить тяжесть наследственных дефектов, а в некоторых случаях осуществить полную коррекцию наследственного заболевания. Высокая пластичность генотипа создает большие возможности для лечения, профилактики наследственных болезней, разработки эффективных медико-педагогических программ обучения, реабилитации и адаптации больных детей.
Литература
1. Дубинин генетика. – М.: Наука. – 1970. – 1486 С.
2. , Шкурат генетики человека. – Учебное пособие. - Ростов н/Д., «Феникс». – 1997. – 368 С.
3. , , Морозов генетики и наследственные нарушения развития детей. - М.: Асадема. – 2003. – 215 С.
4.
Оглавление
Введение…………………………………………………………………………...3
Молекулярные основы наследственности. Строение и функции нуклеиновых кислот………………………………………………………………………………4
Структурно-функциональная единица наследственной информации – ген…12
Организация генома……………………………………………………………..15
Заключение……………………………………………………………………….18